|
| But
precisely! Modern evolutionary theory claims that species do not change gradually, but “abruptly”! |
The idea that species “morph” gradually into other species is an old one, dating to Charles Darwin’s time (mid-19th C.), though not necessarily to Darwin himself.(*) Back then evidence was scarce, so it was impossible to determine the rates at which species change. Moreover, there were some antagonistic theories, such as Lamarck’s,(*) which claimed that individuals (not just species) could acquire novel traits from the environment, and pass them to their offspring. For example, Lamarck thought that the iron smith’s children are strong because the iron smith acquires strong muscles by working daily with the iron and passes this characteristic to his children. Also, Georges Cuvier had supported the theory of catastrophism, a consequence of which concluded that species appear relatively suddenly after catastrophic geological events.(*) It was against this background of a mixture of misunderstanding and confusion that Darwinists claimed that species (not individuals) change gradually by means of natural selection, over the course of thousands or millions of years. Many biologists who argued for evolution after Darwin’s time (early 20th C.) assumed a wrong model, according to which species have some supposed innate tendency to evolve; thus, if nothing happens, it was thought that species will gradually evolve to different species.
This idea, although in line with the Victorian prevailing thought of inherent progress in society, was nonetheless wrong. As was mentioned in the previous article, species “reflect” (in an abstract way) the properties of their environment. If nothing that threatens the survival of a species happens in its environment, the species will live “happily” in its stable niche. But if — as we saw in the example of the aquatic frogs that turned to tree-dwellers — an environmental pressure appears that threatens its survival, then the originally stable and “happy” species might give rise to a number of short-lived transitional species, until one or more of the transitional species find new stable niches (if that happens), and live there “happily” evereafter, seen not as transitional anymore but as stable,(*) until the next environmental pressure, of course. The short-lived transitional species, together with some other reasons that will be explained soon, are the cause of the observed gaps in the fossil record.
Let’s make a drawing of this idea. First, the idea of gradual constant change(*) is shown schematically in the following figure:
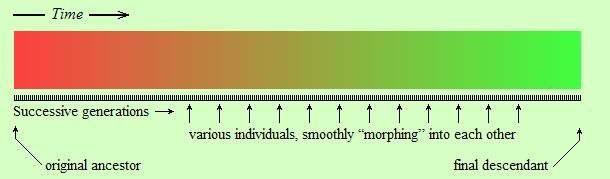
The original ancestor species is represented by individuals of red color in the above figure. Each individual gives birth to another individual, represented by the next little black line under the colored strip. Each descendant differs from its parent (in color in this figure, but in physical traits in reality), but the difference is so tiny that we cannot notice it merely by looking at the parent and the child. However, looking at a sufficiently distant descendant (green color) over a long period of time we can see that there is a definite difference (red vs. green) between original ancestor and distant descendant.
But, as I said, this view is most likely wrong. The following figure seems to be a more accurate representation of reality:
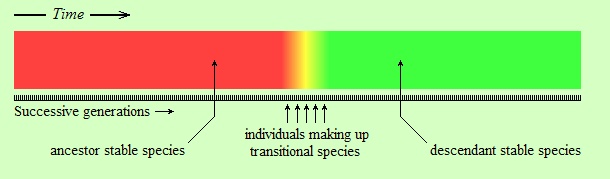
The difference in the above figure is that the change is relatively abrupt. The word “relatively” in this context is important. The individuals shown in orange, yellow, and lime in the figure (labeled as “individuals making up transitional species”) might have lived for several thousand years. However, the entire colored strip from left (red) to right (green) is supposed to have lasted for tens of millions of years, or more. Thus, the transitional species are represented by a very narrow piece of time on the strip. Therein lies the first reason of the fossil record appearing as having gaps. Because, suppose that you can take a few random snapshots within this time interval, represented by the black vertical lines on the following strip (assume each black line is a fossil):
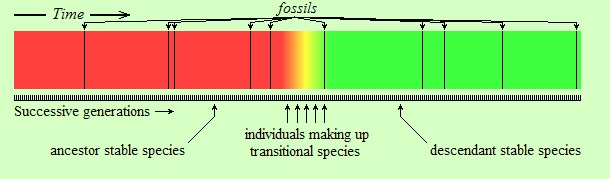
As you see, the snapshots (fossils), being random and sparse, “missed” the transitional species (orange, yellow, and lime) entirely. In reality, the reader should imagine a much narrower interval of time for transitional species than the one shown above, and a sparser set of fossil snapshots. Consider: if the length of the entire strip is, say, 10 million years, and if the length of the transitional period is 50 thousand years (reasonable numbers for evolutionary standards, depending on the kind of living beings considered),(*) then the transitional period would be only five thousandths of the entire length; it would take a width of only about two pixels to represent it in the above figure. Imagine having to “hit” that kind of interval in your sample of fossils. It is in this sense that we say that species evolve to other species “abruptly”: not literally abruptly (not from parent to child, of course, as Lamarck thought), but so fast that when seen from a faraway perspective the transitional period almost vanishes, and the change appears as if it is abrupt.
The above model of abrupt evolutionary change is called the “punctuated equilibrium” model. It says that long periods of little change (relative “equilibrium”) are interrupted (“punctuated”) by short periods of rapid change.(*)
In addition to the above, there is one more reason that contributes to the scarcity of transitional fossils:
| Fossilization is an extremely rare event |
Just because a creature dies does not imply it will become a fossil! Special conditions must be present for fossilization to take place. For example, favorable conditions include the dead creature being covered by the volcanic ash of a nearby volcano that erupted, or by the mud of a lake or river that flooded. But not all creatures live close to an erupting volcano, or by the banks of a river! Even then, such conditions do not guarantee the preservation of the creature as a fossil; they only increase the probabilities that fossilization will take place. Lastly, a creature must contain some hard parts, such as bones, to fossilize; bacteria, jellies, worms, and other soft-bodied creatures, generally do not leave fossils behind. According to Richard Cowen (in History of Life) the majority of animal species living now are unlikely ever to leave fossils, since they are soft-bodied invertebrates such as worms and slugs. Consider that of the more than 30 phyla of living animals, two-thirds of these have never been found as fossils! For all the above reasons, transition from “red” to “green” might happen, as in the previous figures, but if none of the “yellow”, etc., transitional individuals fossilized, then the change went undocumented in the fossil record. No matter how hard we dig, a transitional individual will not be unearthed, because none is there to be found. In that case it is not the paleontologist’s ax, but nature itself that “missed” recording the transition.
And yet, the savvy reader would think, there isn’t only a single instance in which a species evolved to something else; there have been billions of instances of evolution. Missed once, missed twice — well, at some point we should discover a case of transitional fossils. Isn’t that only logical?
Exactly, it is. And that’s precisely what has occurred: in a few lucky cases — bingo! Paleontologists unearthed transitional fossils.
Sometimes, some people — typically non-biologists — deny that these are transitional species. So, if an “orange” fossil is unearthed, they think it’s “red”, because it looks “red” to non-experts; if it’s a “lime” fossil, they’ll categorize it as “green”; and if it’s a “yellow”, then their black-and-white logic forces them to call it either “red” or “green”, whatever makes more sense to their untrained eye. What follows in the rest of this article hopefully will help to clarify such misunderstandings.
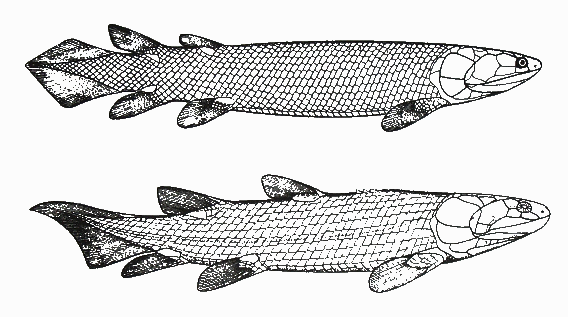
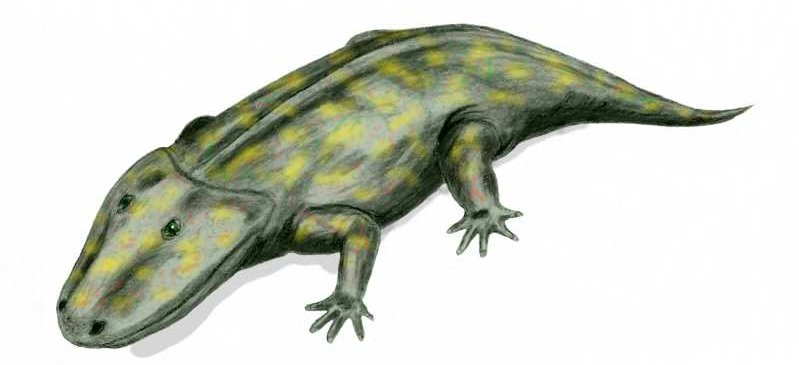
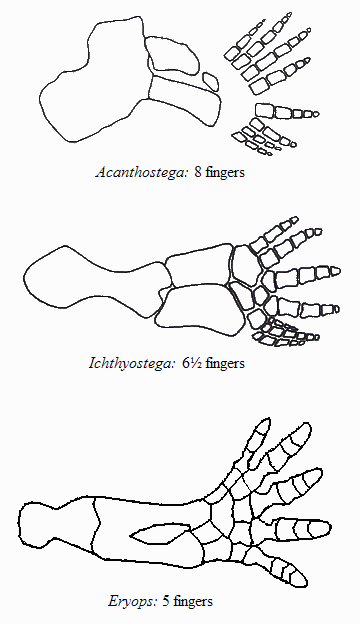
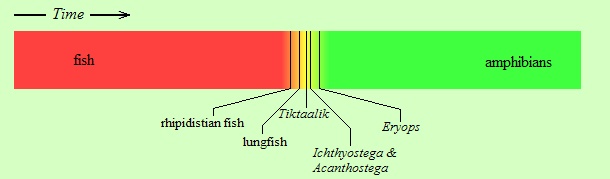
Sometimes it has happened that the transition occurred between so vastly different life-forms that the transitional species that emerged cannot possibly be lumped into either the “red” or the “green” category. Such was the situation when aquatic vertebrates (bony fish), in the course of millions of years, “came out” of the water and “conquered” the land, first yielding literally “fish with legs”, i.e., transitional creatures that disappeared (but will be presented below), then the amphibians, and from them the reptiles, mammals, and birds. Here, “came out of the water” does not mean that a single individual fish acquired legs and started wandering triumphantly on land! It means that, species after species, along some particular evolutionary branch within the tree of evolution, individuals were becoming more adapted to life on land, and less adapted to living in water. Of course, that evolution resembles a tree (rather than a linear chain) means that at the same time other branches of the tree were evolving but not toward moving out of water. The transition that yielded amphibians from fish is very important from an evolutionary perspective, not only because transitional creatures have been found and will be shown below, but also because by studying this transition we learn multiple important principles of evolution. Specifically,
-
we can talk about the notion of adaptive radiation, and
-
we learn that there are usually some “left over” creatures that remind us of such transitions, which survive in specialized niches; in this case, the “left overs” comprise the class of amphibians, i.e., mostly frogs and salamanders.
Hmm... did I say “frogs”? Why don’t we let our friend, the tree frog, who explained to us the story of its ancestors in the previous article, to come back and tell us how its much-much earlier ancestors, the fish, came out of the water. Note that although the story will be recounted by a froggy, it’s not “just a story”, or a legend, but an account of that particular stage of vertebrate evolution that has been amply documented with fossil evidence. Let’s give the podium to the frog now.